
Photo: Joseph A. Sisneros
|
I am a sensory biologist and acoustician interested in the information content of acoustic signals, and how animals perceive sound. I am also interested in the algorithms used by animals to locate a sound producing object solely via hearing. For my PhD dissertation, I am using the acoustically communicating intertidal nesting toadfish, the plainfin midshipman (Porichthys notatus) as a model to address these broad questions. After spending the winter offshore in deep waters (>100 m), midshipman migrate to rocky intertidal beaches during the summer (May-September) for breeding. Type I or “singing” males nest underneath rocks from which they produce nocturnal, long duration advertisement calls (“hums”) to attract females. These hums are the longest contiguous vocalizations in the animal kingdom! Females localize the nests of humming males in near complete darkness and return to deeper water after spawning. Type I males engage in parental care till the eggs hatch and the fry leave the nest. Type II or sneaker males do not produce hums but rather sneak into the nests of type I males and fertilize a fraction of the eggs. Thus, the midshipman is a great model to answer the following questions:
1) What information do type I males convey to females via their mating hums? 2) How do females perceive the hums? 3) How do females hone in on the location of the humming males? |
Internship with Applied Ocean Sciences
I completed a summer internship with Applied Ocean Sciences from June to September 2023, under the mentorship of Dr. Kerri Seger. I worked on two projects:
Propagation models of gharial pops in a river environment
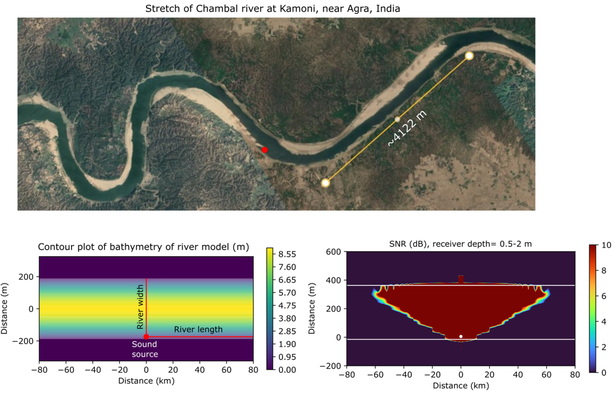
Applied Ocean Sciences received a grant from the Office of Naval Research (ONR) to implement Empirical Mode Decomposition (EMD) algorithms for automating the detection and classification of underwater sounds between 2020 and 2023, referred to hereafter as the "EMD project." Within this project, the EMD algorithm underwent testing with multiple acoustic datasets collected in various aquatic environments worldwide. The algorithm demonstrated its effectiveness in both detecting and classifying acoustic signals, even in conditions with a signal-to-noise ratio (SNR) as low as 0 dB re 1µPa.
One of the objectives of the EMD project was to estimate the detection range of classified signals using acoustic propagation models. I contributed to this goal by estimating the detection range of "pop" sounds produced by male Indian gharials (Gavialis gangeticus) in the Chambal River in northern India. These recorded pop sounds formed a key component of the acoustic datasets used to assess the EMD algorithm's effectiveness. This dataset was graciously provided by Jailabdeen AjjiM, Project Coordinator at the Gharial Ecology Project, Madras Crocodile Bank Trust. To fulfill this task, I employed Applied Ocean Sciences' in-house software known as "Peregrine" to construct parabolic equation propagation models for propagation loss (PL) of the gharial pops. Peregrine, historically used in marine environments, was adapted for a freshwater environment in collaboration with Richard Campbell of AOS. Given the unavailability of high-resolution bathymetric data for the Chambal River at sites where gharial pops were recorded, I created a simplified parabolic bathymetric profile based on the Chambal's dimensions. Bottom sound speed profiles were estimated from soil particle size data, and PL of the peak frequency of the gharial pop was modeled. The modeled sound source was placed at the hydrophone used to record the pops. Signal-to-noise ratios (SNR) for the peak frequency at depths where gharials typically produce pops were calculated using the sonar equation. The horizontal range from the hydrophone at which SNR fell below 0 dB re 1µPa was considered the detection range of gharial pops using the EMD algorithm, following the principle of acoustic reciprocity.
One of the objectives of the EMD project was to estimate the detection range of classified signals using acoustic propagation models. I contributed to this goal by estimating the detection range of "pop" sounds produced by male Indian gharials (Gavialis gangeticus) in the Chambal River in northern India. These recorded pop sounds formed a key component of the acoustic datasets used to assess the EMD algorithm's effectiveness. This dataset was graciously provided by Jailabdeen AjjiM, Project Coordinator at the Gharial Ecology Project, Madras Crocodile Bank Trust. To fulfill this task, I employed Applied Ocean Sciences' in-house software known as "Peregrine" to construct parabolic equation propagation models for propagation loss (PL) of the gharial pops. Peregrine, historically used in marine environments, was adapted for a freshwater environment in collaboration with Richard Campbell of AOS. Given the unavailability of high-resolution bathymetric data for the Chambal River at sites where gharial pops were recorded, I created a simplified parabolic bathymetric profile based on the Chambal's dimensions. Bottom sound speed profiles were estimated from soil particle size data, and PL of the peak frequency of the gharial pop was modeled. The modeled sound source was placed at the hydrophone used to record the pops. Signal-to-noise ratios (SNR) for the peak frequency at depths where gharials typically produce pops were calculated using the sonar equation. The horizontal range from the hydrophone at which SNR fell below 0 dB re 1µPa was considered the detection range of gharial pops using the EMD algorithm, following the principle of acoustic reciprocity.
Detection of underwater sounds using PAMGuard
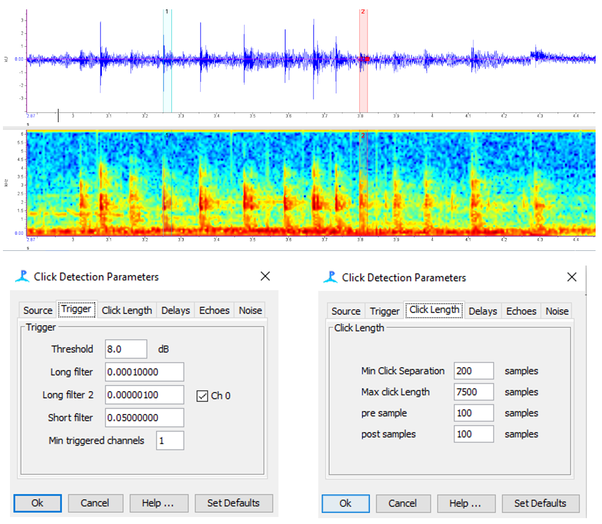
Another objective of the EMD project (See text under the previous heading for more details) was to assess the performance of the EMD process in comparison to the software commonly utilized within the marine bioacoustics community. One of the most widely employed software tools for detecting and classifying sounds from passive acoustic data is PAMGuard. I developed detectors in PAMGuard for the same datasets on which the EMD algorithm was tested. Depending on the nature of the sound, I created either Ishmael energy summation detectors, click detectors, or whistle/moan detectors. Each dataset consisted of multiple audio segments. After constructing the detector in PAMGuard, it was then evaluated on each audio segment, and the detection or absence of the sound signal of interest was documented. True positives, false positives, and true negatives were recorded to compute precision, recall, and accuracy. The precision, recall, and accuracy of the detectors constructed on PAMGuard were subsequently compared to that of the EMD algorithm by Dr. Kerri Seger.
More details about my work at Applied Ocean Sciences can be found in this report for the Office of Naval Research, titled 'Application of an Empirical Mode Decomposition (EMD) Detection and Classification Process to Environments for Naval Monitoring and Detection,' to which I contributed as an author:
More details about my work at Applied Ocean Sciences can be found in this report for the Office of Naval Research, titled 'Application of an Empirical Mode Decomposition (EMD) Detection and Classification Process to Environments for Naval Monitoring and Detection,' to which I contributed as an author:
Dissertation research
I am pursuing my dissertation in the lab of Dr. Joseph Sisneros, University of Washington Seattle, USA.
Honest acoustic signaling in type I male plainfin midshipman
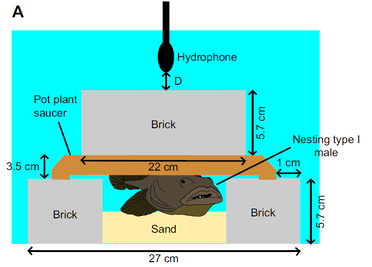
Type I or "singing" male plainfin midshipman produce mating vocalizations called "hums" which constitute the longest known uninterrupted vocalizations in the animal kingdom. The longest recorded hums have been close to 2 hours in duration. Prior to this study, it was unknown whether the hum contains information about the 'quality' or reproductive potential of the type I male. We recorded the hums of type I males in a laboratory setting and found that the loudness of the hum was positively correlated with body size (body mass and standard length). Harmonic frequencies of the hum increased with body condition up to a threshold beyond which the harmonic frequencies remained relatively constant with improvement in body condition. Larger type I males tend to have greater fitness (father more offspring) in the wild. Females preferred to spawn in the nests of larger males in a laboratory setting. Females were also more attracted to louder hums in sound playback experiments. Our study suggests that females could potentially use loudness to choose the nests of larger males with greater reproductive potential. The harmonic frequencies of the hum have previously been shown to linearly increase with temperature. In our study, we demonstrated that if the temperature is maintained constant, the harmonic frequencies are dependent on body condition, with males in poor condition producing lower harmonic frequencies on average. Therefore, hums of type I males serve as honest indicators of size and body condition.
This study has been published in the Journal of Experimental Biology: Balebail and Sisneros, 2022.
This study has been published in the Journal of Experimental Biology: Balebail and Sisneros, 2022.
Perception of harmonic structure of mating calls in female midshipman

Pitch perception plays an important role in human communication. A key feature of pitch perception is the ability to recognize the harmonic structure of an acoustic signal. The ability to perceive harmonic structure has been documented in mammals, birds, and even some species of frogs. The ecological significance of possessing this perceptual ability, however, remains poorly understood. Perceiving harmonic structure might be adaptive for female plainfin midshipman. Type I or "singing" males produce long duration mating calls ('hums') to attract females for spawning. The hum is like a musical note, possessing a harmonic structure with a fundamental frequency (f0) of 80-120 Hz and several higher harmonic frequencies (all being integral multiples of f0). Midshipman breed in shallow water (< 5 m deep), which functions as a natural high pass filter, allowing only the higher harmonics to propagate long distances. Therefore, perceiving harmonic structure of a hum even when f0 is absent might be crucial for mate call detection and localization in female plainfin midshipman.
We hypothesized that female midshipman can perceive the hum’s harmonic structure even in the absence of f0, and that they possess the ability to discriminate a periodic hum from an aperiodic hum lacking a harmonic structure. To test these hypotheses, we performed two-choice acoustic trapping experiments in the natural rocky intertidal breeding environment of midshipman at Seal Rock, Brinnon, WA, USA during the summers of 2021 and 2022. Acoustic stimuli were relayed during the night through underwater speakers contained inside two fish traps placed 2-4 m apart. The following acoustic stimuli were synthetically generated: an average hum, a hum with diminished f0, and an acoustic signal containing the same f0 as a hum but with scrambled higher harmonic frequencies (aperiodic hum). The number of females captured overnight in each trap represented the attractiveness of the acoustic stimulus. Overall, our results are suggest that the fundamental frequency (f0) is both necessary and sufficient to attract female midshipman, and the harmonic note-like structure is not an important factor in mate attraction. Therefore this species might be highly vulnerable to low frequency anthropogenic sounds in the ocean, which might mask the fundamental frequency. Whether the midshipman or other fish species have the ability to detect the harmonic structure of sounds in general remains an open question. To the best of my knowledge, this study represents the first attempt to study mate call perception in the natural environment in fishes. I am in the process of writing up these results for publication.
We hypothesized that female midshipman can perceive the hum’s harmonic structure even in the absence of f0, and that they possess the ability to discriminate a periodic hum from an aperiodic hum lacking a harmonic structure. To test these hypotheses, we performed two-choice acoustic trapping experiments in the natural rocky intertidal breeding environment of midshipman at Seal Rock, Brinnon, WA, USA during the summers of 2021 and 2022. Acoustic stimuli were relayed during the night through underwater speakers contained inside two fish traps placed 2-4 m apart. The following acoustic stimuli were synthetically generated: an average hum, a hum with diminished f0, and an acoustic signal containing the same f0 as a hum but with scrambled higher harmonic frequencies (aperiodic hum). The number of females captured overnight in each trap represented the attractiveness of the acoustic stimulus. Overall, our results are suggest that the fundamental frequency (f0) is both necessary and sufficient to attract female midshipman, and the harmonic note-like structure is not an important factor in mate attraction. Therefore this species might be highly vulnerable to low frequency anthropogenic sounds in the ocean, which might mask the fundamental frequency. Whether the midshipman or other fish species have the ability to detect the harmonic structure of sounds in general remains an open question. To the best of my knowledge, this study represents the first attempt to study mate call perception in the natural environment in fishes. I am in the process of writing up these results for publication.
Role of the swim bladder in directional hearing
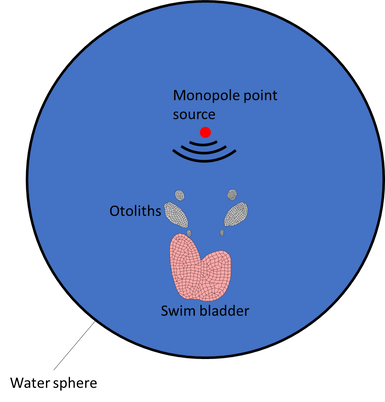
Sound is a mechanical disturbance passing through an elastic medium which causes particles in the medium to vibrate (termed as "particle motion") and also causes local fluctuations in pressure (called "sound pressure"). The ears of terrestrial vertebrates detect sound pressure whereas fishes primarily detect sound via particle motion. When the sound source is a monopole and close to the animal, the position of the source can be computed by triangulating the particle motion vectors at two different points in space. Fish auditory systems have been shown to use vector-sensing to localize sound sources in the near field (At distances much smaller than the wavelength). However, when the sound source is located far away from a fish, there is a 180 degree ambiguity in determining the direction of the sound source via particle motion alone. How do fishes resolve this 180 degree ambiguity in directional hearing? The gas filled swim bladders that many fishes possess allow the detection of sound pressure. When a sound passes through a fish, the hair cells (auditory receptors) in the inner ears are directly activated by particle motion created by the sound source. Due to changes in pressure due to the passage of the sound, the swim bladder expands and contracts, functioning like a secondary sound source, generating particle motion which stimulates the inner ears, thus potentially informing fishes about sound pressure. The phase model has long been proposed as a mechanism to resolve the 180 degree ambiguity in directional hearing in fishes. It predicts that fishes can determine the direction of the sound source by computing the phase relation between sound pressure and particle motion, which changes predictably depending on the direction of the sound source. However, this hypothesis remains largely untested. I am using a numerical modelling technique (finite element analysis, FEA) on a specialized FEA software called COMSOL to test this hypothesis. I am generating 3d models inspired by the the actual shapes of the swim bladder and inner ear bones (otoliths) of female plainfin midshipman, which have been shown to localize the sounds of calling type I males with high accuracy. I am currently simulating how the inner ear bones move in response to monopole point sources or plane wave sources, in the presence and absence of the swim bladder. To begin with, I am using a simplified model, enclosing the swim bladder and otoliths in a water sphere, assuming that the acoustic properties of fish tissues to be the same as water. In future, these simple models can be expanded to incorporate material properties of bones and even soft tissues, and the activation profiles of the hair cell fields located close to the otoliths.
Natural underwater sounds as sources of information and noise for fishes

In aquatic bioacoustics, sounds other than conspecific communication sounds have generally been viewed as background noise interfering with the detection of communication signals. However, many fish species do not vocalize but still possess acute hearing, suggesting that fish hearing may have evolved, in part, to detect and extract information from the “ambient sounds” present in the environment. We wrote a review in The Effects of Noise on Aquatic Life, Principles and Practical Considerations highlighting some of the major sources of natural ambient sound and discusses how they function as sources of biologically relevant information. Fishes have been known to obtain information about larval settlement sites, spawning events, food sources, and approaching predators from natural ambient sound. A survey of literature also revealed instances where natural ambient sounds function as noise. Ambient sound can mask hearing and some vocalizing fishes have been known to either produce louder vocalizations or change vocalization rates to cope with increases in ambient sound levels. Several species living in shallow water habitats vocalize at frequencies that coincide with a dip in acoustic energy in the local ambient sound spectrum. As we wrote the review, we realized that data on the behavioral and physiological responses of fishes to natural ambient sounds remains scarce. This information could be important for understanding how anthropogenic sounds affect the survival and reproduction of fishes.
The review titled "Natural Ambient Sounds as Sources of Biologically Relevant Information and Noise for Fishes" has been published in The Effects of Noise on Aquatic Life, Principles and Practical Considerations.
The pdf can be downloaded here:
The review titled "Natural Ambient Sounds as Sources of Biologically Relevant Information and Noise for Fishes" has been published in The Effects of Noise on Aquatic Life, Principles and Practical Considerations.
The pdf can be downloaded here:
Past research
Landing maneuvers of houseflies (Musca domestica) on vertical and inverted surfaces (Master's project)
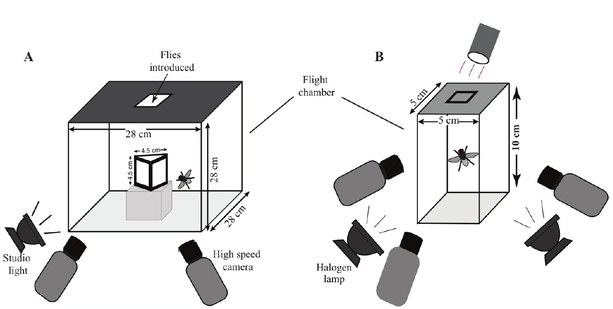
I did my master's research in the lab of Dr. Sanjay Sane, National Centre for biological Sciences, Bengaluru, India.
Insects can land on substrates having different orientations and textures. High speed videos of insect landings have revealed that landing maneuvers are complex behaviors which can be decomposed into sequences of modules, like body-deceleration, leg-extension, and body rotations. Despite the complexity of the maneuver, landings are performed quite quickly, within a fraction of a second. Prior to our study, it was unknown if insects use similar or different 'rules' or algorithms to land on substrates oriented in different directions. We conducted a series of experiments in which houseflies (Musca domestica) were lured to land on vertical or inverted surfaces. Their landing trajectories were recorded with multiple highspeed cameras filming at 3000-4000 fps. We observed that well-controlled landings occurred on both surfaces when the distance at which flies began slowing down was proportional to the component of flight velocity perpendicular to the direction of the landing surface. The ratio of distance from the landing surface to velocity at the onset of deceleration (called 'tau' or 'time to collision') was conserved, despite substantial differences in the mechanics of vertical and inverted landings. Flies which did not begin slowing down at this value of tau bumped their head against the landing surface, suggesting that these landings were less controlled. Flies also extended their legs before touchdown. Unlike deceleration, leg-extension was independent of approach velocity or how far the flies were from the landing surface. Thus, the robust reflexive visual initiation of deceleration is independent of substrate orientation, and combines with a more variable initiation of leg-extension. Together, these combinations of behaviors likely enable flies to land in a versatile manner on substrates of various orientations. The simple landing algorithms that we have uncovered could be utilized to guide landings in human made flying objects.
This study was published in Plos One: Balebail et al., 2019.
Insects can land on substrates having different orientations and textures. High speed videos of insect landings have revealed that landing maneuvers are complex behaviors which can be decomposed into sequences of modules, like body-deceleration, leg-extension, and body rotations. Despite the complexity of the maneuver, landings are performed quite quickly, within a fraction of a second. Prior to our study, it was unknown if insects use similar or different 'rules' or algorithms to land on substrates oriented in different directions. We conducted a series of experiments in which houseflies (Musca domestica) were lured to land on vertical or inverted surfaces. Their landing trajectories were recorded with multiple highspeed cameras filming at 3000-4000 fps. We observed that well-controlled landings occurred on both surfaces when the distance at which flies began slowing down was proportional to the component of flight velocity perpendicular to the direction of the landing surface. The ratio of distance from the landing surface to velocity at the onset of deceleration (called 'tau' or 'time to collision') was conserved, despite substantial differences in the mechanics of vertical and inverted landings. Flies which did not begin slowing down at this value of tau bumped their head against the landing surface, suggesting that these landings were less controlled. Flies also extended their legs before touchdown. Unlike deceleration, leg-extension was independent of approach velocity or how far the flies were from the landing surface. Thus, the robust reflexive visual initiation of deceleration is independent of substrate orientation, and combines with a more variable initiation of leg-extension. Together, these combinations of behaviors likely enable flies to land in a versatile manner on substrates of various orientations. The simple landing algorithms that we have uncovered could be utilized to guide landings in human made flying objects.
This study was published in Plos One: Balebail et al., 2019.